


Introducción
El encanecimiento del cabello se correlaciona con el envejecimiento cronológico y ocurre en todos los individuos, independientemente del sexo y la raza. Se considera que el 50 % de los individuos de raza caucásica tienen al menos el 50 % del cabello canoso a la edad de 50 años1. Aunque las canicies prematuras suelen aparecer sin una enfermedad subyacente, se han asociado con anemia perniciosa, hipotiroidismo, osteopenia2 y con varios síndromes raros. Existen casos anecdóticos de reversión de las canas tras inyecciones de epinefrina3, tras simpatectomía4, por vasoconstricción y por exposición continua a bajas temperaturas.
En la piel humana existen al menos dos poblaciones diferentes de melanocitos: los epidérmicos y los del folículo piloso (FP)5,6. La melanogénesis del FP está relacionada con el estadio de crecimiento del ciclo capilar, mientras que la melanogenesis epidérmica es continua7 e influenciable por la radiación ultravioleta. Los melanocitos del FP pueden dividirse en tres subpoblaciones diferentes. La primera, localizada en el bulbo folicular, sólo expresa la proteína relacionada con la tirosinasa 2 (TRP2), no prolifera y, presumiblemente representa a los melanocitos stem cells del FP. La segunda, localizada en la vaina folicular externa del FP, expresa TRP2 y en menor medida la proteína relacionada con la tirosinasa 1 (TRP1), presenta actividad proliferativa durante el anagen temprano e intermedio y representa a los melanocitos diferenciados. La tercera, localizada en la matriz del pelo sobre la papila dérmica, expresa TRP1, TRP2 y tirosinasa, prolifera sólo durante el anagen intermedio, produce melanina durante el anagen intermedio y tardío y progresivamente desaparece en la fase de catagen8. Las dos últimas subpoblaciones descritas expresan c-kit y tienen dependencia de la señal stem cell factor (SCF)/c-kit durante la fase de anagen9.
El mejor modelo para estudiar el ciclo capilar es un modelo de ratón, altamente estandarizado, denominado C57BL/6, donde la depilación manual induce el crecimiento sincrónico del pelo terminal10. Por el color de la piel de los ratones tras la depilación, podemos distinguir fácilmente la fase de telogen (rosado) de la fase de anagen (gris a negro). Se trata de un modelo muy utilizado donde están perfectamente definidas la duración de cada fase del ciclo capilar y su correlación clínico-patológica10. Durante el período entre telogen y anagen los melanocitos en reposo proliferan, se diferencian y migran dentro del FP de forma sincronizada para regenerar y construir un nuevo bulbo folicular.
La hormona tiroidea es esencial en la homeostasis de las células derivadas de la cresta neural, de forma que un hipotiroidismo congénito produce un daño neurológico grave. En la piel la hormona tiroidea ejerce una función conocida en la diferenciación epidérmica, en el incremento de la respuesta local a los factores de crecimiento, en la fisiología de las glándulas sebáceas, ecrinas y apocrinas, y en el crecimiento del cabello.
En este trabajo, tomando como base los hallazgos clínicos observados en dos pacientes, discutimos el posible papel de la hormona tiroidea en el oscurecimiento del pelo canoso. Además, investigamos el efecto de la triiodo-tironina (T3) sobre el FP en un modelo animal de ratón, de cara a definir el efecto de esta hormona sobre la función melanocitaria. Finalmente corroboramos in vitro, sobre unidades foliculares humanas, un estímulo de la T3 en el crecimiento del tallo piloso. Partiendo de estos resultados, y revisando la literatura, formulamos varias hipótesis sobre el papel de los melanocitos no sólo en la repigmentación sino en toda la homeostasis folicular.
Material y métodos
Casos clínicos
Paciente 1
Un varón de 63 años con el pelo encanecido progresivamente a lo largo de los años, y una micosis fungoide (estadio IA) de un año de evolución en tratamiento con PUVA (3 sesiones/semana), debutó con un coma mixedematoso de causa desconocida. Acudió a la consulta acompañado por un familiar para realizar una sesión de PUVA con un bajo nivel de conciencia, respiración irregular, inflamación y rubefacción facial y de ambas manos, y edema periorbitario. La tensión arterial era de 110/60 mmHg, el pulso de 56/minuto y la temperatura de 35° C. En la analítica realizada destacaba una T3 libre de 0,4 pg/ml (normal 1,5-4,3), una T4 libre de 1,8 pmol/ml (normal 10,5-25,0) y una hormona estimulante del tiroides (TSH) de 102 mU/l (normal 0,45-7,0). El paciente fue ingresado en la Unidad de Cuidados Intensivos (UCI) durante 10 días, donde recibió medidas de soporte y tratamiento intravenoso con 0,5 mg de L-tiroxina. La evolución fue satisfactoria, continuando el tratamiento por vía oral con 0,1 mg de L-tiroxina, que permitió el regreso a casa en un mes. Tras la salida de la UCI se apreció una completa repigmentación de todas las canas previas del cuero cabelludo, que ha permanecido estable durante 2 años de seguimiento. Su enfermedad de base no experimentó cambios significativos y continuó requiriendo tratamiento específico. El paciente no recibió ninguna otra medicación durante el ingreso.
Paciente 2
Se trata de un varón de 51 años afecto de síndrome de Cowden con antecedente de carcinoma de tiroides, tratado mediante cirugía y tratamiento hormonal sustitutivo con hormona tiroidea. Progresivamente con el paso del tiempo había ido sufriendo un encanecimiento de su cabello. Dos años más tarde desarrolló un linfoma no hodgkiniano de amígdala y ganglios cervicales que fue tratado con quimioterapia y radioterapia (dosis total de 36 Gy). En ese momento el 60 % de su cabello era blanco. Después de completar 4 ciclos de quimioterapia (CHOP [ciclofosfamida, doxorubicina, vincristina, prednisona y bleomicina]) acudió a la consulta con un estado de nerviosismo, ansiedad, insomnio y pérdida de 15 kg de peso, siendo diagnosticado de hipertiroidismo iatrogénico debido a una descompensación del tratamiento sustitutivo (0,2 mg/día de L-tiroxina). La analítica realizada mostró unos niveles de T3 libre de 1,62 pg/ml, una T4 libre de 34,4 pmol/l y una TSH indetectable (< 0,1 mU/l). Durante este período presentó una repigmentación total de su cabello. Los niveles de T4 libre fueron normales tres meses después de un tratamiento diario con 0,15 mg de L-tiroxina. Durante un período de seguimiento de 3 años ha persistido el pelo pigmentado.
Los dos pacientes recibieron dosis elevadas de hormona tiroidea, uno debido a un coma mixedematoso y el otro por descompensación de su enfermedad. Aunque no pueden descartarse totalmente otras causas de reversión de las canas como, por ejemplo, la evolución de un linfoma cutáneo o el tratamiento con PUVA en el primer paciente11, o el tratamiento con quimioterapia en el segundo12, atribuir a la hormona tiroidea la pigmentación del cabello parece la hipótesis más plausible y razonable. En el primer paciente la repigmentación no es atribuible a los psoralenos ni a su asociación con luz ultravioleta A, ya que este tratamiento lo venía realizando desde meses antes sin ningún efecto sobre el color del cabello.
Ratones
El cuidado y la manipulación de los animales se realizó de acuerdo con el protocolo habitual. Se usaron ratones hembra C57BL/6 (Jackson Labs, Bar Harbor, ME, USA) de 6 semanas de edad, con un peso aproximado de unos 20 g. Una vez aclimatados y tras una semana en observación, los animales fueron divididos en 2 grupos de 5 y el dorso fue depilado con cera fría10. El experimento fue aprobado por la comisión de investigación correspondiente.
La triiodo-tironina fue obtenida de Sigma (Barcelona, España). Utilizamos el modelo de ratón C57BL/6, estandarizado para el estudio del ciclo capilar10. Concretamente, la cera fría fue aplicada en el dorso de los ratones de 7 semanas, apreciándose de forma uniforme tras la depilación, un color rosado de la piel indicativo de que todos los folículos estaban en la fase de telogen. Con la ayuda de una micropipeta, se administraron tópicamente 0,5 μg de T3 disueltos en 20 μl de etanol en el dorso de todos los ratones de uno de los grupos, mientras que en el otro se utilizaron 20 μl de etanol como control. La aplicación fue diaria y el tiempo del estudio fue de 10 días.
Cultivo de unidades foliculares
Tras el consentimiento informado, mediante la disección con microscopio, se obtuvieron unidades foliculares sobrantes en fase de anagen de la región occipital de dos mujeres (30 y 38 años) que fueron sometidas a trasplante capilar. Las unidades foliculares se cultivaron en medio de Williams E (Gibco, Madrid, España) con glutamina, 1 % de suero de ternera fetal, insulina (10 ng/ml), transferrina (10 μg/ml), hidrocortisona (10 ng/ml) y selenito sódico (10 ng/ml). Para evitar la sobreinfección, el medio de cultivo se trató con penicilina/estreptomicina (50 U/ml-50 3 ug/ml) y fungizona (2,5 μg/ml)13. Se hicieron 2 grupos, uno control y otro con el mismo medio de cultivo añadiendo 100 nM de T3. Las unidades foliculares se mantuvieron en semiflotación en placas de 24 pocillos, en estufa a una temperatura uniforme de 37° C, con una atmósfera de 95 % de aire y 5 % de CO2. El medio se cambió cada 72 horas. Se realizaron fotografías con cámara digital bajo los mismos parámetros, utilizando un microscopio invertido binocular, al iniciar el cultivo (basal), a las 72 y a las 144 horas.
Resultados
Ratones
A la edad de 7 semanas todos los ratones C57BL/6 presentaban un característico color rosado en la piel visible tras la depilación, indicativo de la fase de telogen (fig. 1). Cuando los ratones entraron en anagen comenzó el recrecimiento del pelo, caracterizado por una progresiva pigmentación del dorso de gris a negro. Tras 5 días de tratamiento con 0,5 μg/día de T3 el 100 % de los animales testados entró en anagen (fig. 2). En el grupo control se apreció un incremento de la pigmentación sugestivo del inicio del anagen- a partir del día 9, pero no se observó en el 100 % de los ratones hasta el día 15.

Figura 1. Ratones hembra C57BL/6 de 7 semanas depilados con cera fría por tracción, previa anestesia superficial con ketamina a dosis convencional. Se aprecia una coloración rosada uniforme de la piel alopécica, característica de una fase sincrónica de telogen.
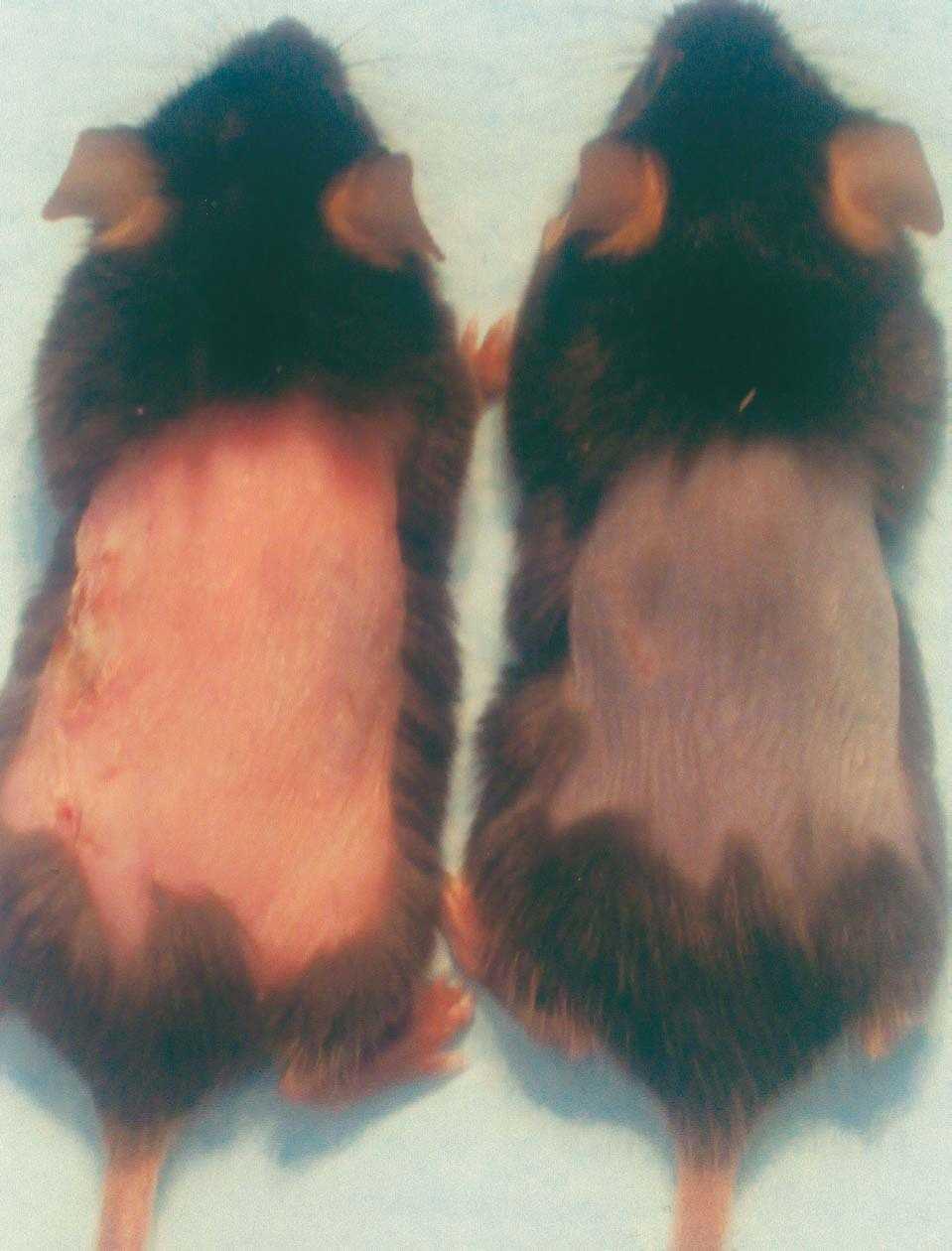
Figura 2. A los 5 días de tratamiento se observa un cambio de coloración de la piel en el ratón tratado con T3 tópica (derecha), frente al ratón control (izquierda), sugestivo de la entrada sincrónica en anagen.
Tras 8 días de tratamiento se realizó un estudio histológico para confirmar la inducción de la fase de anagen. La piel, estirada y adherida a un papel fotográfico para evitar pliegues, fue fijada en formol, lavada con etanol y embebida en parafina según la técnica habitual. Se realizaron tinciones con hematoxilina-eosina y tricrómico de Masson (fig. 3). Todos los experimentos se repitieron dos veces.
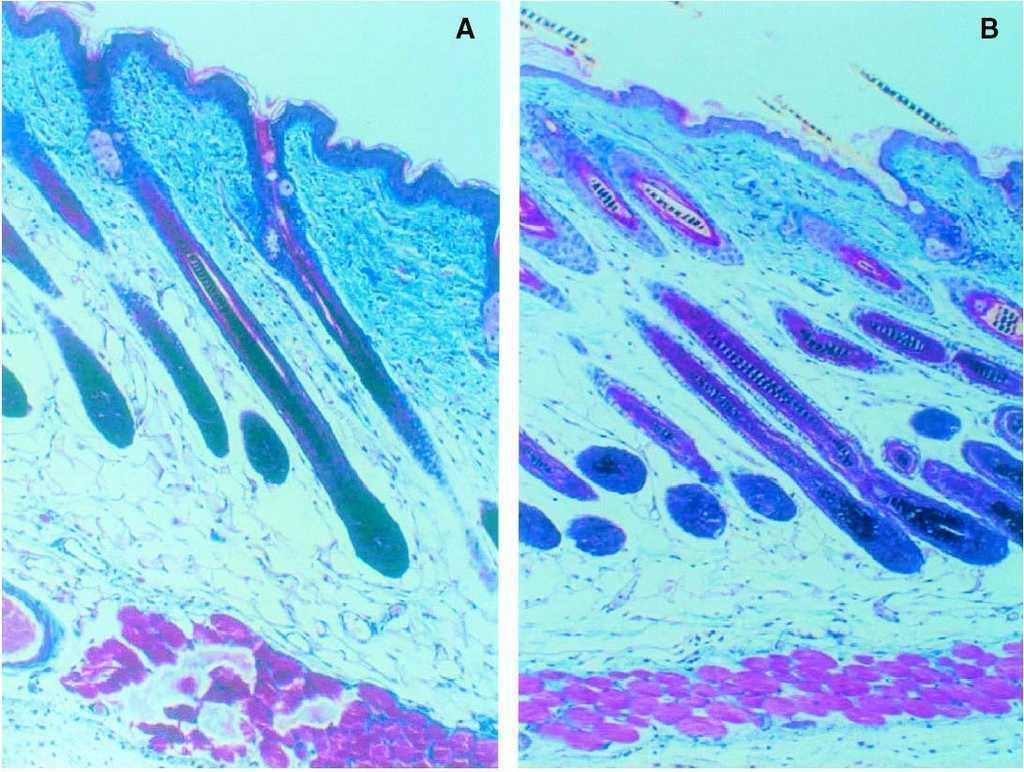
Figura 3. Tinción de tricrómico de Masson sobre la biopsia realizada el día 10. (A) En el grupo control se observan folículos en fase de anagen (IV), donde la punta del nuevo tallo piloso todavía no alcanza la epidermis (×100). (B) En el grupo tratado con T3 tópica se aprecia una mayor densidad de folículos y un anagen más avanzado (VI), con pelos emergiendo a través de la epidermis (×100).
Cultivos
De cada donante se utilizaron 8 unidades foliculares por grupo. Los resultados fueron muy similares en ambos casos. A las 72 horas en uno de los experimentos se apreció un crecimiento significativo en 6 de las 8 unidades foliculares tratadas con T3, mientras que sólo hubo un crecimiento en 3 de las 8 unidades foliculares del grupo control (fig. 4). Durante este tiempo la arquitectura folicular se conservó completamente. Las mediciones se realizaron sobre papel tras ampliar uniformemente las fotografías realizadas. La media de crecimiento para el grupo tratado con 100 nM de T3 fue de 1,2 ± 0,05 mm (media ± desviación estándar) a las 72 horas, frente a los 0,65 ± 0,05 mm del grupo control. A la dosis de 200 nM de T3 no se incrementaron las diferencias (datos no mostrados).
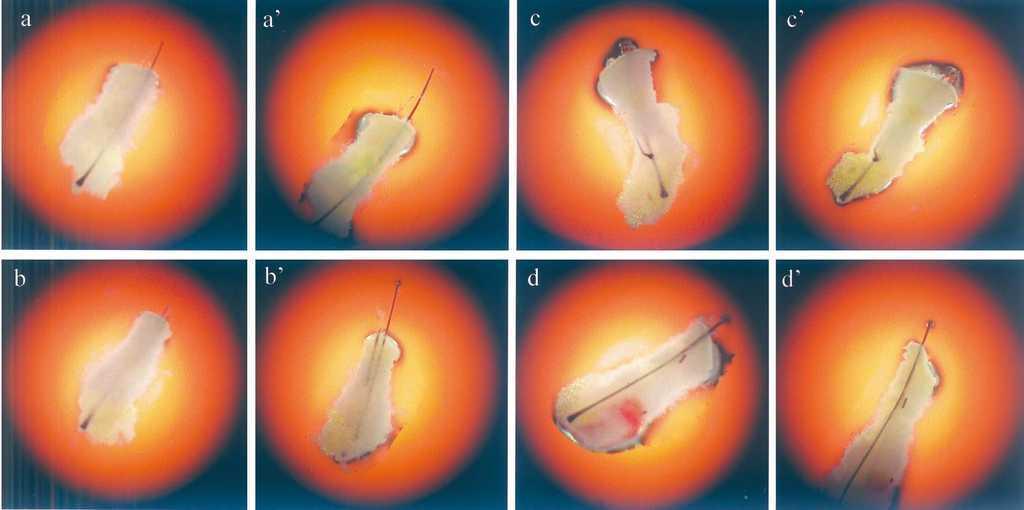
Figura 4. Unidades foliculares en situación basal (a-d) y a las 72 horas (a9 -d9) del cultivo in vitro. Se aprecia un crecimiento significativo del tallo piloso en las muestras tratadas con 100 nM de T3 (a, b) respecto a las muestras control (c, d). Fotografías representativas de todos los experimentos.
Discusión
El encanecimiento del pelo es debido a una alteración (ralentización o fracaso) en la repoblación del nuevo bulbo piloso en anagen por melanocitos «frescos» derivados de la vaina folicular externa14. La expresión de tirosinasa y la actividad melanogénica sólo son detectables en la subpoblación de melanocitos foliculares cercana a la papila dérmica. Esta estructura, la papila dérmica, secreta factores de crecimiento tipo SCF y factor de crecimiento hepatocitario que, probablemente, estén involucrados en la transición de melanocitos del FP a formas completamente diferenciadas productoras de melanina9.
Varios trabajos de la literatura han demostrado que el encanecimiento capilar no es un proceso irreversible. Hay evidencias aisladas que muestran la repigmentación del cabello después de la radioterapia por cáncer o después de procesos inflamatorios como el eczema eritrodérmico, la candidiasis erosiva del cuero cabelludo15, el carbunco16 y las forunculosis17. También se ha comunicado un oscurecimiento temporal del pelo después de la administración de altas dosis de ácido para-aminobenzoico18 y después del uso de fármacos como verapamilo19, tamoxifeno20, ciclosporina21, etretinato22, levodopa23, latanoprost24 y arsénico25. Asimismo se han publicado casos anecdóticos de repigmentación del pelo canoso tras la exposición a rayos X, después del uso del PUVA en un paciente con síndrome de Sézary11, en dos pacientes con enfermedad celíaca26 y en dos pacientes con porfiria cutánea tarda27.
La piel es una diana bien conocida de las hormonas tiroideas. La hipodensidad pilosa en el cuero cabelludo, la caída del tercio externo de las cejas y la disminución del vello corporal aparecen frecuentemente asociados al hipotiroidismo y son debidos a un incremento de la proporción de folículos en telogen respecto a los folículos en anagen28. Esta situación puede corregirse tras la administración de L-tiroxina, implicando así a la hormona tiroidea en el ciclo capilar y en el crecimiento del cabello humano29. El FP tiene receptores TR-α y TR-β para la hormona tiroidea, especialmente TR-β30, 31, que han sido detectados en las células de la vaina folicular externa, en las células de la papila dérmica y en los fibroblastos que rodean la envoltura externa del folículo. Además, estudios in vitro con células de la vaina folicular externa y la papila dérmica han demostrado que la hormona tiroidea puede tener un efecto directo, a través de sus receptores específicos, sobre el crecimiento del FP32. Nuestro trabajo demuestra cómo la aplicación tópica de T3 acelera la entrada del pelo en anagen en un modelo murino estandarizado. En un estudio in vivo, con ratones SKH-1 (ratones sin pelo por una mutación genética) y ratas, se apreció que la administración tópica de T3 en liposomas aumentaba significativamente el número y el tamaño de los pelos frente a un grupo control y otro grupo tratado a bajas dosis, sin evaluar su influencia específica en alguna fase concreta del ciclo capilar33. Más interesante es un trabajo reciente que demuestra una influencia variable de la hormona tiroidea sobre la piel en función de la administración (tópica o intraperitoneal). Concretamente, los ratones que recibieron T3 intraperitoneal presentaban una epidermis un 10 % más adelgazada y menor densidad de folículos (48 % menos); por el contrario, a los ratones a los que se les administró la T3 tópicamente tenían la epidermis un 78 % más engrosada y una mayor densidad de folículos (160 % más). Estos datos sugieren que aunque la T3 estimula la proliferación queratinocitaria, también influye sobre los fibroblastos dérmicos para la liberación de factores inhibidores de la proliferación34. Por otra parte, la administración de T3 a perros hipotiroideos favorece la entrada del pelo en anagen, en comparación con perros eutiroideos o perros hipotiroideos no tratados35.
Estudios in vitro sobre unidades foliculares han demostrado cómo el factor transformador de crecimiento beta (TGF-β), los ésteres del forbol (TPA), el factor básico de crecimiento de fibroblastos (bFGF) y el suero de ternera fetal reducen el crecimiento de los tallos pilosos y tienen un papel importante en la inducción de la fase de catagen del ciclo capilar13, 36. Este efecto, de causa desconocida, podría explicarse por el hecho de que ambos (TGF-β37 y bFGF38) estimulan la expresión y producción de c-kit. Los melanocitos pueden regular el ciclo capilar, la pigmentación y la entrada/salida en anagen-catagen mediante la expresión de c-kit39. Si se inhibe la expresión de c-kit con anticuerpos anti-TGF-β o con inhibidores selectivos del TGF-β, por ejemplo, se pierde la pigmentación del tallo piloso y se alarga la fase de anagen40. Todas estas sustancias (TGF-β, TPA, bFGF, etc.) tienen en común el favorecer la proliferación y diferenciación de los melanocitos en cultivos puros. En este sentido, la hormona tiroidea también es necesaria en el cultivo de melanocitos, al igual que ocurre con otras células derivadas de la cresta neural. Así, inferimos que los melanocitos podrían ser una de las dianas celulares de todas estas sustancias (incluyendo la hormona tiroidea) cuando actúan sobre los FP. De esta manera se puede especular sobre el papel que desempeña la hormona tiroidea en la homeostasis melanocitaria, tanto sobre las células névicas (crecimiento de nevus)41 como sobre los melanocitos epidérmicos en un proceso tan frecuente como el vitíligo, donde se ha demostrado una disfunción tiroidea hasta en el 30 % de los casos de nueva aparición42. Concretamente, como ya se ha comentado, las hormonas tiroideas participan en la maduración de las células derivadas de la cresta neural a través de la regulación de receptores β2-adrenérgicos. Así, por ejemplo, la maduración y diferenciación de los astrocitos que se produce tras la administración de la hormona tiroidea, es mediada por receptores β2-adrenérgicos43. Sobre el cultivo de melanocitos, algunos fármacos adrenérgicos al estimular estos receptores β2, incrementando el AMPc, favorecen la melanogénesis y la proliferación celular44. Por el contrario, el uso de fármacos bloqueadores β puede favorecer el comienzo de un cuadro de vitíligo en pacientes predispuestos45. Finalmente, algunos preparados que han sido eficaces en el tratamiento de determinados casos de vitíligo contenían hormona tiroidea46.
En un plano mucho más teórico, y obviando la interacción de otros múltiples factores neuro-endocrinos, podemos inferir que la disminución de la hormona tiroidea afecta a los melanocitos, igual que ocurre con el resto de las células derivadas de la cresta neural. En ocasiones el blanqueamiento brusco del cabello puede ir precedido por una experiencia estresante o trastornos neuropsiquiátricos, en muchas ocasiones relacionados con una disminución de la hormona tiroidea o un aumento de sus necesidades. Popularmente se acepta que la canicie precoz en un corto espacio de tiempo es fruto de las preocupaciones y de un estado tensional. Así, un susto, un episodio de estrés, una ansiedad mantenida, etc. (situaciones en las que histórica y culturalmente está muy bien documentada la aparición de una alopecia areata o un encanecimiento brusco del cuero cabelludo) pueden producir un aumento brusco de las necesidades de T3. Ante la imposibilidad de una mayor producción es posible que las primeras células que se resientan sean los melanocitos foliculares. La melanogénesis epidérmica es constante y quizá con un menor «requerimiento hormonal», mientras que la melanogénesis folicular es cíclica, más puntual y posiblemente con un mayor «requerimiento hormonal».
Los melanocitos del FP alcanzan su máxima capacidad proliferativa durante el anagen temprano e intermedio, fases en las que asocian la expresión de c-kit en su superficie. La pérdida de pigmentación de los tallos pilosos en las canas no se debe a una pérdida completa de todos los melanocitos foliculares; de hecho se ha demostrado la presencia de melanocitos dopa-negativos y otros muchos marcadores específicos negativos en la vaina folicular externa de cabellos blancos seniles47. Este hallazgo deja abierta la posibilidad futura de estimular la migración y diferenciación de los melanocitos para favorecer la repigmentación de los FP canosos.
Evidentemente la hormona tiroidea puede influir sobre otras estirpes celulares, no estudiadas ni discutidas en este trabajo, que están presentes en la piel normal como son los queratinocitos y los fibroblastos. Además, un punto esencial para verificar la hipótesis anterior, que en nuestro caso no hemos podido realizar debido a la imposibilidad de obtener unidades foliculares despigmentadas, es valorar su posible repigmentación in vitro mediante el cultivo selectivo con T3. Aunque no es habitual la presencia de canas en la región occipital (zona donante de pacientes sometidos a trasplante capilar) esperamos en un futuro próximo poder disponer de estas unidades foliculares, para confirmar los hallazgos descritos.
En resumen:
1.La hormona tiroidea estimula el crecimiento de los tallos pilosos, in vitro, sobre cultivo de unidades foliculares.
2.Tras la aplicación tópica de T3 en ratones C57BL/6, los folículos en fase de telogen entran más precozmente en anagen.
3.En seres humanos, la hormona tiroidea puede revertir las canas, repigmentando el pelo terminal. Los melanocitos foliculares podrían ser la diana de al menos estas dos últimas acciones.
La base de este trabajo estriba en unos hallazgos clínicos sobre el papel de la hormona tiroidea en la repigmentación folicular; posteriormente, en los modelos habituales, hemos demostrado otras acciones, más o menos conocidas, de la T3 en la regulación del ciclo capilar. Llegados a este punto establecemos una posible conexión sobre el papel de los melanocitos no sólo en la repigmentación sino en toda la homeostasis folicular. Así, parece lógico que estas células de morfología neuronal, derivadas de la cresta neural, puedan desempeñar un papel esencial en la regulación del ciclo capilar y no sólo limitarse a proporcionar un color al cabello. En el día a día apreciamos cómo el pelo canoso de la barba y del cuero cabelludo, es más grueso y más resistente a la caída que el pelo pigmentado. La manipulación farmacológica, utilizando los melanocitos como «nuevas dianas», podría abrir otras líneas de investigación en el crecimiento capilar, diferentes de la dirigida a bloquear la miniaturización producida por las hormonas sexuales, que es la única que parece tener vigencia hoy en día.
Conflicto de intereses
Declaramos no tener ningún conflicto de intereses.
Correspondencia:
Pedro Redondo.
Departamento de Dermatología.
Clínica Universitaria de Navarra.
31008 Pamplona. España.
predondo@unav.es
Aceptado el 25 de abril de 2007.


